
Age is a significant deciding factor in many behavioral tests. In certain cases, behaviors that are present early in life may not be present later, while the inverse is also sometimes true. Regarding other types of animal behavior, especially in the social and cognitive domains, behaviors may simply be different at different ages. Here, we will see several examples of both of these possibilities.
Ultrasonic vocalizations
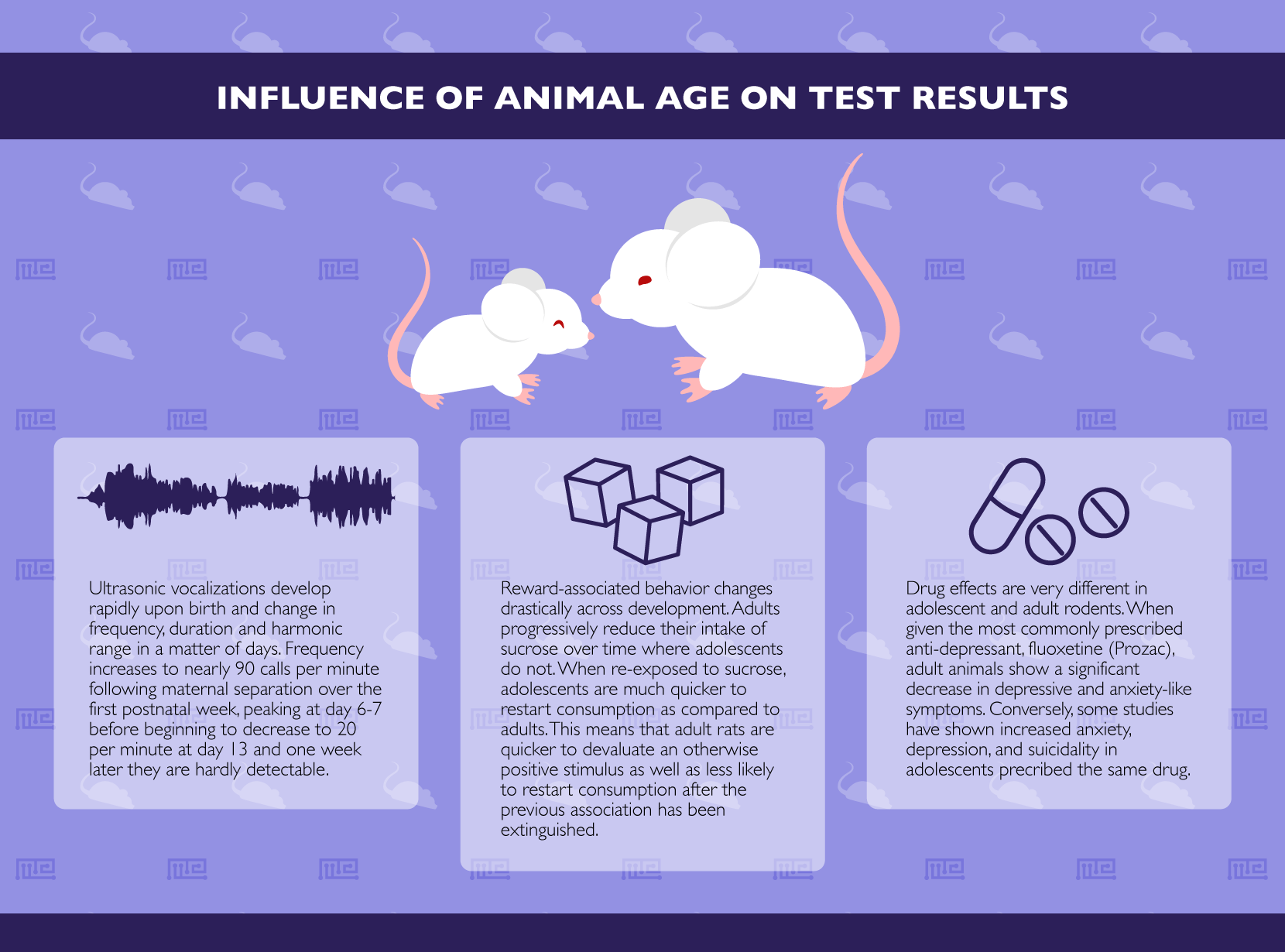
Amongst the behavioral analyses unique to early development, ultrasonic vocalizations (USV) are perhaps the most well-studied. USV develop rapidly upon birth and change both in their frequency (the number of calls), duration and harmonic range in a matter of days.[1] While they are detectable within the first days of life, their frequency continues to increase to nearly 90 calls per minute following maternal separation over the first postnatal week. This frequency peaks at postnatal day 6-7 before beginning to decrease.[2] By postnatal day 13, the number of calls made is reduced to less than 20 per minute and one week later they are hardly detectable. USV made by adult animals are much more rarely studied and are thought to represent entirely different behaviors. Specifically, while calls made by young, developing rodents are distress signals induced by maternal separation, those made by adult rodents are thought to indicate either predator stress or appetitive (reward-like) perception, depending on the harmonic range.[3] Thus, USV are a prime example of a behavior which changes both in its magnitude and meaning as a function of developmental age.
Reward behavior
One behavioral domain which changes quite drastically across development is reward-associated behavior. This is particularly important and interesting because one of the major reasons for testing reward behavior in animals is to model addiction, which is itself an age-dependent phenomenon in humans.[4] As an example, a robust 2014 study was designed to examine the effects of age on the entire reward behavior process from acquisition to devaluation and eventually, extinction and reacquisition.[5] Using a pavlovian conditioning protocol which paired an auditory stimulus (a tone) with a reward (sucrose delivery), the authors studied this time-course in both male and female rats beginning either during adolescence or adulthood.
While adolescents and adults were equally likely to develop a positive association between the tone and the reward, significant differences were found in the later stages of the testing. Specifically, by allowing free access to the sucrose and then initiating an extinction protocol (wherein the solution was provided as before but without any sucrose), the researchers decreased the value of this reward. They found that while adults progressively reduced their intake over time, adolescents did not. Then, when the animals were re-exposed to the original paradigm (termed “reacquisition”), the adolescents were much quicker to restart consumption as compared to adults. Thus, in the case of rewarding behavior, adult rats are quicker to devaluate an otherwise positive stimulus as well as less likely to restart their consumption after the previous association has been extinguished. Importantly, similar results have been shown with other rewarding stimuli, such as drugs of abuse.[6]
Drug effects: Anti-depressants
The most commonly prescribed anti-depressant, fluoxetine (Prozac), has very different effects in adolescent and adult rodents.[7] Predictably, adult animals show a significant decrease in depressive and anxiety-like symptoms when given fluoxetine. Indeed, countless studies have shown that this is the case, leading to fluoxetine being one of the most widely prescribed drugs on the planet.
Conversely, a 2009 study found that when administered in clinically relevant doses to juvenile mice (both Swiss Webster and C57Bl/6 strains), fluoxetine-induced, rather than diminished, anxiety.[8] Specifically, using the open field and elevated plus mazes, juvenile mice were found to spend more time in hidden or obscured areas of the mazes, indicating heightened stress. These findings are in line with reports of adverse outcomes in adolescents prescribed the drug which includes increased anxiety, depression, and suicidality.[9]
Conclusions
There are many additional examples of behavioral tests whose outcomes are altered as a function of age. As gonadal hormones develop over the periods of adolescence and adulthood, social behavior changes in tandem.[10] Models of neurodegeneration show no symptomology until late-adulthood in most cases,[11] while performance in a variety of memory-associated tasks shows an inverted-U curve wherein their effects peak during early adulthood and begin to decline with significant aging.[12] Stressful experiences imbue substantially different behavioral outcomes depending on the age during which they occur.[13] Surely, there is no shortage of evidence that age is a significant factor in behavioral testing. Thus, it is important for all researchers to carefully consider the age of the animals that they are testing when designing any behavioral task and understand that this can be a decisive factor in what tests are most appropriate and how the outcome data are to be interpreted.
References
- Branchi, I., Santucci, D., & Alleva, E. (2001). Ultrasonic vocalisation emitted by infant rodents: a tool for assessment of neurobehavioural development. Behavioural Brain Research, 125(1–2), 49–56.
- Elwood, R. W., & Keeling, F. (1982). Temporal organization of ultrasonic vocalizations in infant mice. Developmental Psychobiology, 15(3), 221–227.
- Brudzynski, S. M. (2009). Communication of adult rats by ultrasonic vocalization: biological, sociobiological, and neuroscience approaches. ILAR Journal, 50(1), 43–50.
- Galvan, A. (2010). Adolescent Development of the Reward System. Frontiers in Human Neuroscience, 4.
- Hammerslag, L. R., & Gulley, J. M. (2014). Age and Sex Differences in Reward Behavior in Adolescent and Adult Rats. Developmental Psychobiology, 56(4), 611–621.
- Anker, J. J., & Carroll, M. E. (2010). Reinstatement of cocaine seeking induced by drugs, cues, and stress in adolescent and adult rats. Psychopharmacology, 208(2), 211–222.
- Olivier, J. D. A., Blom, T., Arentsen, T., & Homberg, J. R. (2011). The age-dependent effects of selective serotonin reuptake inhibitors in humans and rodents: A review. Progress in Neuro-Psychopharmacology & Biological Psychiatry, 35(6), 1400–1408.
- Oh, J., Zupan, B., Gross, S., & Toth, M. (2009). Paradoxical anxiogenic response of juvenile mice to fluoxetine. Neuropsychopharmacology: Official Publication of the American College of Neuropsychopharmacology, 34(10), 2197–2207.
- March, J., Silva, S., Petrycki, S., Curry, J., Wells, K., Fairbank, J., … Treatment for Adolescents With Depression Study (TADS) Team. (2004). Fluoxetine, cognitive-behavioral therapy, and their combination for adolescents with depression: Treatment for Adolescents With Depression Study (TADS) randomized controlled trial. JAMA, 292(7), 807–820.
- Bell, M. R. (2018). Comparing Postnatal Development of Gonadal Hormones and Associated Social Behaviors in Rats, Mice, and Humans. Endocrinology, 159(7), 2596–2613.
- Creed, R. B., & Goldberg, M. S. (2018). New Developments in Genetic rat models of Parkinson’s Disease. Movement Disorders: Official Journal of the Movement Disorder Society, 33(5), 717–729.
- Tan, H. M., Wills, T. J., & Cacucci, F. (2017). The development of spatial and memory circuits in the rat. Wiley Interdisciplinary Reviews. Cognitive Science, 8(3).
- Urban, K. R., & Valentino, R. J. (2017). Age- and Sex-Dependent Impact of Repeated Social Stress on Intrinsic and Synaptic Excitability of the Rat Prefrontal Cortex. Cerebral Cortex (New York, N.Y.: 1991), 27(1), 244–253.