
Effective navigation is a necessary skill for survival. In order to find food, avoid predators, and mate, all members of the Animal Kingdom need to know where they are going. The capacity to learn locations and remember routes in the air and water relies on various measurable neurological functions. This article will introduce a beginner’s understanding of those functions, otherwise known as “spatial learning.”
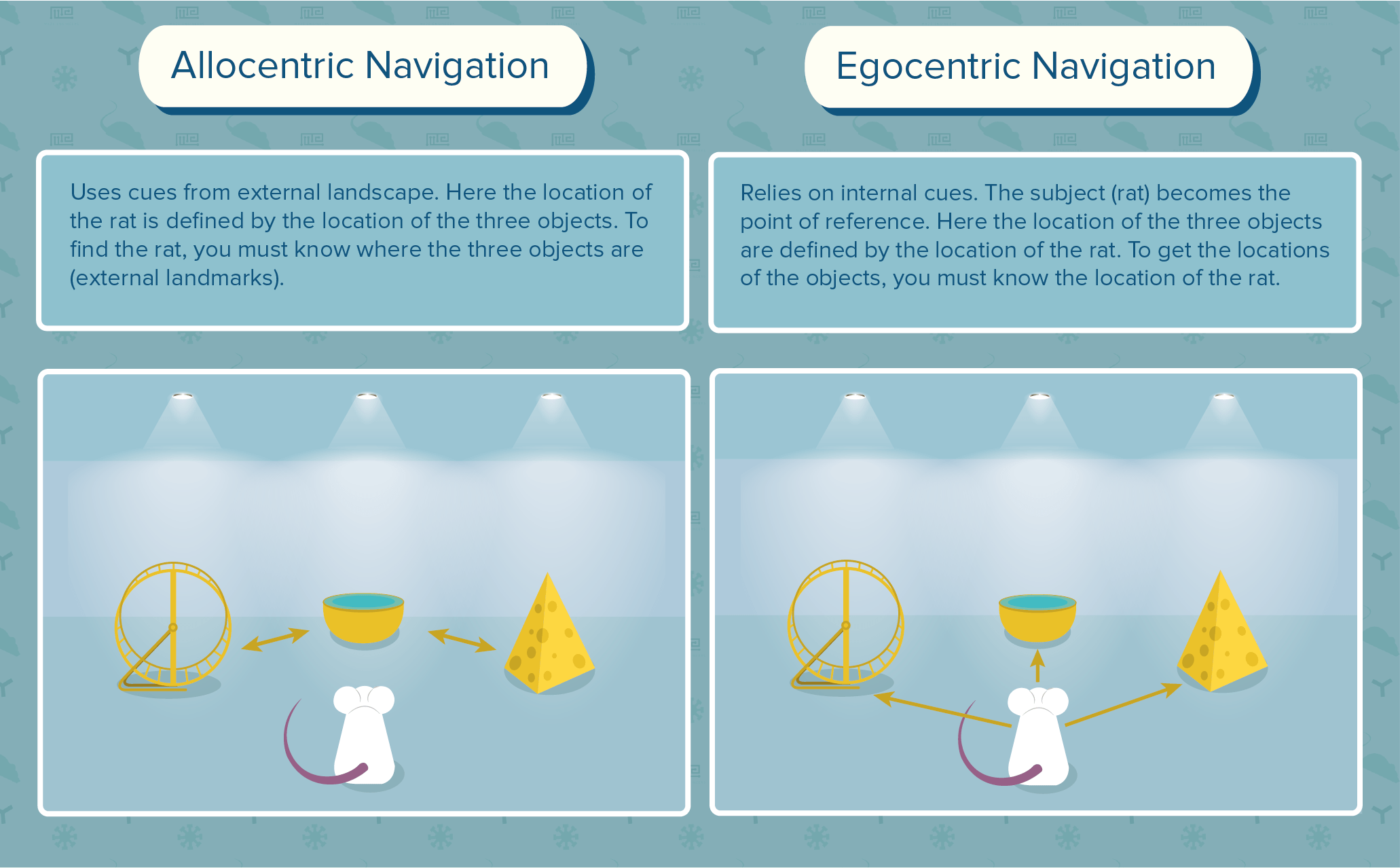
Allocentric vs. Egocentric Navigation
Navigation relies on signposts to indicate directional changes and landmarks. Allocentric navigation, also called “way-finding” or “spatial navigation,” uses cues from external landscape (north, south, east, west), while egocentric navigation relies on internal signals, such as limb movement and turn sequence (to the left, next to, right-hand). For example, imagine giving directions to a friend. If you say, “Go north on I-65,” that would be allocentric navigation, while,”turn left at the McDonalds,” is egocentric.
Egocentric navigation is subdivided into route-based navigation and path integration. Route-based navigation relies on rules, such as turning at specific signposts, and taking a certain number of steps. This has been measured in humans by asking subjects to walk in a circle, then retrace their steps while blindfolded. Compared to the control subjects, patients with perturbations in the striatum are less able to retrace the circle (Paquette et al. 2011)[1]. With repetition, memorized patterns become habitual, often referred to as implicit, or procedural, memory. In humans, this is typically more automatic and less conscious. Think of the route you take to work every day, then getting a different job. You may find yourself on the way to your previous job, and late to your current one!
The second subdivision of egocentric navigation, called path integration, requires vector addition: allowing the organism to find a more direct path without the need to retrace steps. This has been assessed in humans by leading subjects blindfolded around an open space, then asking them to point to their origin and estimate distance in between. Those with damage in the hippocampus are able to complete this task just as well as the control subjects (Shrager et al. 2008)[2], while those with injury to the temporal lobe are unable to complete the task (Buzsaki and Moser 2013)[3]. This proves that navigational systems rely on different neural networks; which should, and can be tested independently despite the overlaps.
Navigation within the Brain
Allocentric navigation is mediated within the hippocampus and entorhinal cortex. The hippocampus is a key structure in forming cognitive maps (O’Keefe and Nadal 1978)[4], and damage in this area results in impaired memory and spatial learning (Brandeis et al. 1989; Burgess et al. 2002; Buzsaki and Moser 2013; McNamara and Skelton 1993; Moser et al. 1998;Penner and Mizumori 2012; Suh et al. 2011; Whitlock et al. 2006)[5][6][7][8][9][10][11][12]. Cells within the hippocampus respond to features in the landscape, creating a neural map of the environment which is then monitored as an organism moves.
The entorhinal cortex tracks direction of movement, while border cells react to boundaries within the environment. This network of specialized cells keeps track of the surrounding environment. In one study, scientists monitored the entorhinal cortex using rats in a Morris Water Maze (MWM). Interestingly, firing rates progressively increased in this network of cells during hidden platform trials, while a visible platform did not trigger the same response (Hollup et al. 2001)[13].
Principles of Navigational Assessment
The key distinctions between allocentric and egocentric navigation are navigational cues. Allocentric navigation uses distal cues, whereas egocentric navigation uses proximal and internal cues. When it comes to experimental testing, an open pool of water removes proximal cues. In human tests, “cue deprivation’ is achieved using blindfolds or total darkness.
One challenge in assessing navigational methods is creating a substantial environment. Subjects should be able to choose their cues during the initial test; it should later be determined which cues were dominant, and how different treatments affect certain memory systems. Another challenge is ensuring the results reflect navigational learning, not performance factors (such as motivation). It is required that all test subjects have equal abilities as it pertains to the experiment; for example, if testing rats in a water maze, be sure that no rat is better or worse at swimming than other rats in the same test.
Assessment Methods
During trials, progressive behaviors are exhibited: thigmotaxis (swimming around the perimeter), swimming further from the edge, then weaving and circling until a submerged platform is found.
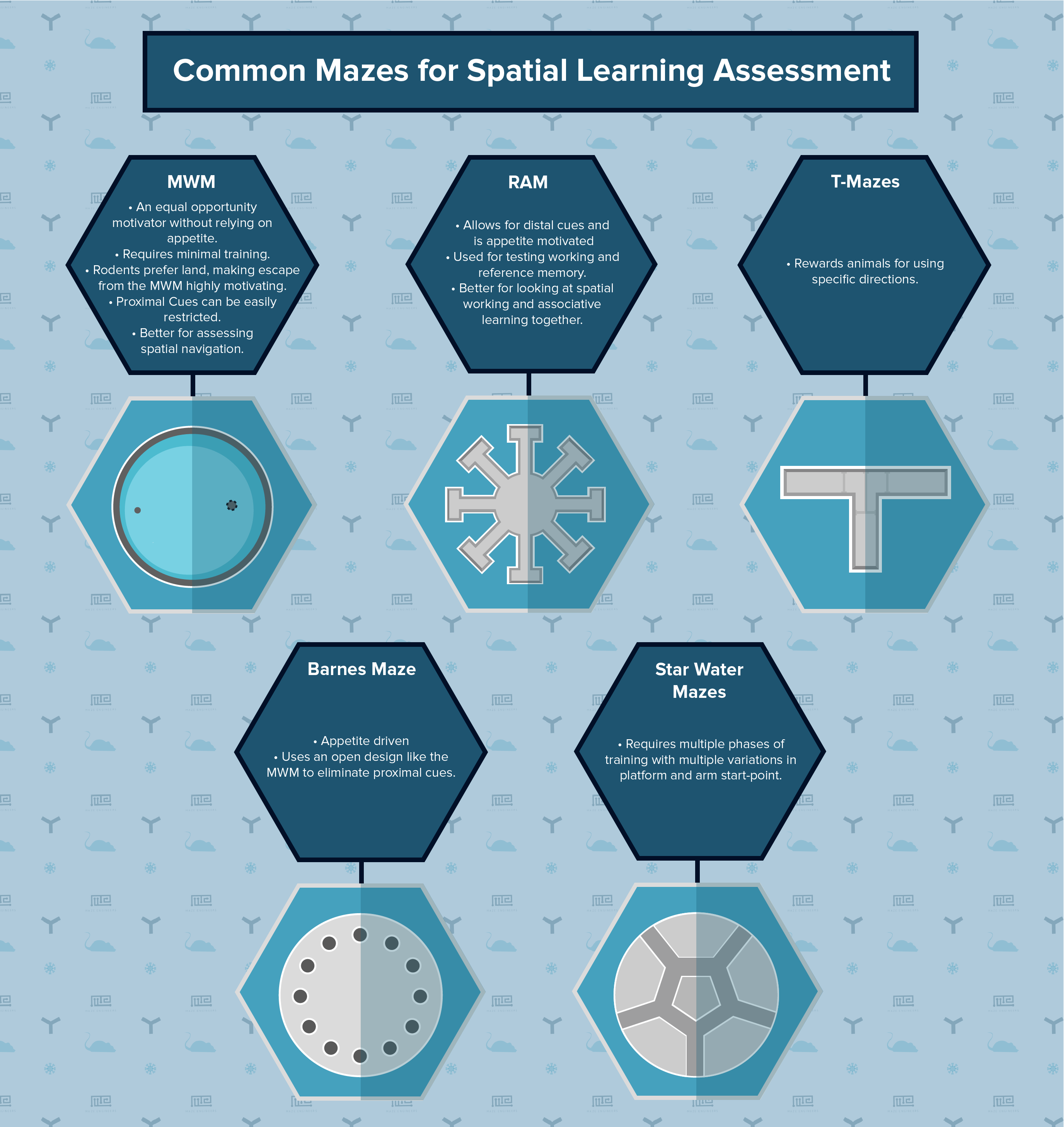
Radial Arm Maze and Morris Water Maze
Spatial learning as well as allocentric navigation can be assessed with a variety of tasks, including the Morris Water Maze or Radial-Arm Maze (RAM). The RAM has been largely superseded by the MWM because it ineffectively removed proximal cues, a problem for all corridor mazes.
The Morris Water Maze is a simple circular pool. It forces distal cues with variations for platform location and visibility, which helps assess navigational learning. MWM research has demonstrated that rodents develop systematic and symmetrical search patterns after repeated exposure. Distal cues are sufficient for learning and developing a simple strategy, which require proximal cues for further optimization.

The Radial Arm Maze allows for distal cues. It is appetite motivated, testing working and reference memory. Variation in arm number increases difficulty and enhances result reliability. The RAM requires food restriction and training in the form of reward exposure. Short-term memory is assessed by placing rewards at the end of the arms, which are re-baited for the next trial – ensuring only short term spatial memory is assessed.
The reliance of food motivation requires standardization of hunger with control groups – difficult in pharmacological trials. Learning is much slower in the RAM compared to the MWM, creating a protracted learning curve. Deviations are clear, and can make interpretation of RAM data a bit more challenging.
The Morris Water Maze is better for assessing spatial navigation, and the Radial Arm Maze is better for looking at spatial working and associative learning together. A hybrid swimming version of the RAM (i.e., radial-arm water mazes [RWM]) removes some motivation challenges and can be used as a quick test of reference memory.
Other Mazes for Spatial Learning Assessment
- T-Mazes: Rewards animals for using specific directions. Creates differentiation between positional and spatial learning by rotating the maze to see if turns are related to cues or habits (although this is typically a maze for rewarded and spotaneous alternation, it has been used in spatial learning in some cases).
- Barnes Mazes: Appetite driven and uses an open design like the MWM to eliminate proximal cues. Target nest box holes are rewarded over multiple days with several trials per day.
- Star Water Mazes: Require animals to swim around a central pentagon before selecting an arm. Multiple phases of training with multiple variations in platform and arm start-point require vector skills to select efficient routes.
Features and advantages of the MWM
- An equal-opportunity motivator without relying on appetite. Assessment of swimming speed is necessary to equalize motor skills and motivation.
- The most widely used test of learning and spatial memory because rodents prefer land environments, making escape from the MWM highly motivating based on the aspect of water.
- Proximal cues can be easily restricted with uniform pool design. Visible platforms provide proximal cues, while curtains remove distal cues.
- 100% completion (in rats and most mice strains) removes bias in appetite driven tests, and nearly 100% task mastery creates quality data sets.
- Performance differences are non-cognitive and non-spatial (thigmotaxis, time in goal quadrant, escape, and floating time), all of which can be adjusted for.
- Requires minimal training due to natural swimming ability and high motivation, creating trial efficiency.
- Characteristics in inbred mice can induce persistent thigmotaxis or floating, instead of searching. Behavioral deficits are also seen in appetite or shock trials.
- The MWM requires adaptions and new platform positions to assess working memory.
Stress Reduction
Water mazes are criticized for undue stress levels, however, food restriction and novel environments also illicit stress; even minimally, stress is required to motivate learning. Typically, memory improves under stress, making it difficult to determine which conditions are negatively stressful for the animals. Isolation adds stress, which slows maze learning (single week isolation has limited impact). Handling increases learning speed, while inescapable stressors reduce learning.
If animals fail to find the platform, guiding or lifting prevents discouragement and development of defeat behaviors. Measuring corticosterone levels in untested versus MWM-tested rats has shown increased plasma levels (30% elevation immediately after the trial, reducing to control levels after 60 minutes). Conducting multiple trials back-to-back can induce a drop in body temperature, especially in mice, making interval periods essential to maintain low stress levels.
Experimental Flexibility and Adaptability of the MWM
- The variation in visible and hidden platforms make the MWM highly adaptable. Specific, working memory versions of the maze are available (Morris 1984;Stewart and Morris 1993)[14][15].
- Use of dual targets and variation in start location allows for discrimination between hippocampal and striatal learning; rats with striatal lesions cannot learn the task, while those with lesions in the hippocampus can.
- Inserting structures into the pool helps poorly performing, inbred strains of mice learn faster; however, structures have little benefit for normal performers and may impact results.
Due to variations in apparatus and testing procedures, researchers are advised to review Morris’ work: foundations (Morris 1981)[16] and methods (Morris 1984; Stewart and Morris 1993)[17][18] in conjunction with methodological improvements (Vorhees and Williams 2006; Wenk 2004)[19][20].
Pool and Platform Size
Pool size was virtually doubled by Morris during his original research. Since then, little systematic study of this variable has been discovered. Faster learning is seen in smaller pools, but if the pool is too small, the task is too easy (Mactutus and Booze 1994)[21].
Pools that are too large could also impact results, but successful learning assessments have been recorded in pools of up to 244-cm (Williams et al. 2014)[22] – a larger boundary has not yet been established. A small pool is typically used for adult mice, which may be beneficial; there is evidence that mice cannot learn in a 210-cm pool (Schaefer et al. 2011)[23]. In a comparison between 122-cm and 152-cm diameter pools where wild-type C57BL/6J mice were used, the mice in the smaller pool found the platform faster on day 1, but on days 2 to 5 and beyond, both groups showed similar rates of improvement.
Search Area to Target Ratio
Target size also impacts task difficulty, and relative ratio of ‘pool-to-target’ allows systematic study relative to task difficulty. A 10cm platform in a 122-cm tank creates a ratio of 149:1 – conditions in which many mouse strains learn well (a 210-cm pool with a 10cm platform shows almost no learning in mice) (Schaefer et al. 2011)[24]. Pool-to-platform ratio can be used to determine task difficulty, but some ratios do not seem to be adequately navigable by mice, unless they are subject to an acceptable ratio first and it is gradually altered.
Rats, on the other hand, learn well even in the higher ratios (larger pools and smaller platforms). Neonatal exposure to methamphetamine has a dramatic difference on performance when compared to controls (Williams et al. 2003b, 2004)[25][26]. Lowering the platform size in stages, however, produced good learning curves in both the methamphetamine exposed and control rats (Vorhees et al. 2008)[27], demonstrating pool-to-platform ratio as moderated by prior experience.
Cues, Platforms and Starting Location
Learning proficiency is drastically changed using distal cues. Rats have a minimal threshold of required cues in allocentric navigation: a minimum of two separate distal cues, in addition to the pool edge (Maurer and Derivaz 2000)[28]. A study in which the starting location moves demonstrates rats with hippocampal lesions use relative position for navigation (Morris et al. 1990, Hamilton et al. 2007)[29][30] and use distal and directional cues together.
Cued exposure before hidden trials reduces problems associated with learning subordinate task skills, such as staying on the platform or searching away from the wall. This has significant value in mice trials, as they are prone to behaviors that reduce learning (thigmotaxis, floating or not recognizing the platform). Trials with cues have positive skill transfer with a modest flattening of the learning curve in just a single day’s exposure, and 6 trials.
Dividing the pool into four cardinal zones creates different start locations for the typical four trials per day. The variation in start locations creates a saw-tooth pattern due to platform proximity, but learning curves must be smooth in order to eliminate distance variance (Morris 1981; Vorhees et al. 1995, Vorhees and Williams 2006)[31][32][33].
Phases and Trial Type
Adding a reversal phase (moving the platform to an opposing quadrant) highlights unseen deficits in simple acquisition phases. Reversal deficits are seen in subjects with hippocampal lesions or receptors blockades, as the animals are unable to ‘give up’ – behaviour otherwise learned through multiple attempts to locate the acquisition platform (Whishaw and Tomie 1997)[34]. This could be the result of spatial learning problems, or interference effects from inflexibility rather than cognitive remapping. If pre-training or cued trails are used before testing, NMDA inhibition is prevalent, eliminating acquisition, but not reversal deficits (Hoh et al. 1999)[35].
Probe trials, a.k.a.”memory trials,” remove the platform where subjects spend their time searching. If the location has been learned previously, the animal will have a preference for the platform’s quadrant. Assessing reference versus working memory relies on sufficient time between trials, otherwise animals use short-term memory instead of consolidated memory. In order to accurately measure reference memory, 24 hours must pass after platform trials (Baldi et al. 2005)[36].
Other factors to be considered regarding Spatial Learning tests
Water temperature: Learning is impaired by significant alternations in core body temperature (Rauch et al. 1989a; Rauch et al.1989b)[37][38]. Aged rats may require warming between trials, but a water temperature of 20–22oC typically works well for both rats and mice.
Inter-trial intervals: Distributed trials increase learning and memory performance, whereas massed trials reduce it (Commins et al. 2003)[39]. Data suggests that spaced trials improve acquisition in rats, but the impact is less significant in mice, and has little impact on reversal or shift trials.
Sex differences: Male rodents demonstrate faster learning in the MWM and better allocentric navigation than females (Hamilton et al. 2007; Vorhees et al. 2008)[40][41]. However, females consistently outperform males in egocentric navigation (Vorhees et al. 2008)[42]. Data from both RAM and MWM tests highlight male navigational performance (Jonasson 2005)[43].
Age differences: Spatial learning is developed after cued learning, therefore younger rats learn the MWM, but require more cues and a smaller pool. Younger animals are less able to find hidden platforms. While egocentric navigation develops pre-weaning, spatial navigational skills come later.
Odor trials: While possible in water mazes, limited or unpublished data is found using pathfinder animals and thorough cleaning between trials. The nature of water mazes minimizes the impact of odor compared to dry mazes.
Rearing and housing: Handled, socialized rodents perform better in mazes. Highly enriched environments improve learning capacity, including maze learning (Tees et al. 1990)[44]. Minimal enrichment facilitates improved maze learning (Vorhees et al. 2008)[45].
Experimental design: study reliability relies on adequate sample size, randomization, blinding, and control of litter effects – this applies to animal studies generally and specifically maze trails.
Sample size: Small samples produce false positives, even if difference seems clear and significant. A sample size of at least 10 is advised. Ideally, 15 to 25 animals per test group is preferred (due to learning variability, a larger sample size is advised for mice).
Litter effects: Overlapping genetic and epigenetic factors with experience make litter mates more similar than rodents from another litter, comparable with fraternal twins. Variance clustering within litters needs to be considered during statistical modeling.
Randomization: Ineffective randomization leads to problems with reproducibility. Arbitrary selection is insufficient. Using identification numbers, in addition to a random number generator is a simple way of replicating selection.
Blinding: Significant bias is introduced when experimenters are aware of treatment type. Inadvertent bias damages results. Documented blinding is crucial.
Conclusion
The MWM has multiple advantages in assessing allocentric navigation in rodents. Some stress is induced, but trials that reinforce learning with food also require stressful situations due to required food deprivation pre-trial. Water mazes are also less stressful than shock methods, and swimming mazes motivate learning and memory with minimal training and high levels of proficiency. Crucially, MWM’s can assess the navigational skills essential for survival and compare them to memory systems in humans.
Reference
[1] Paquette C, Franzen E, Jones GM, Horak FB. 2011. Walking in circles: navigation deficits from Parkinson’s disease but not from cerebellar ataxia.Neuroscience 190:177–183.
[2] Shrager Y, Kirwan CB, Squire LR. 2008. Neural basis of the cognitive map: path integration does not require hippocampus or entorhinal cortex. Proc Natl Acad Sci U S A 105:12034–12038.
[3] Buzsaki G, Moser EI. 2013. Memory, navigation and theta rhythm in the hippocampal-entorhinal system. Nat Neurosci 16:130–138
[4] O’Keefe J, Nadal L. 1978. The Hippocampus as a Cognitive Map. Oxford: Oxford University Press
[5] Brandeis R, Brandys Y, Yehuda S. 1989. The use of the Morris water maze in the study of memory and learning. Int J Neurosci 48:29–69.
[6] Burgess N, Maguire EA, O’Keefe J. 2002. The human hippocampus and spatial and episodic memory. Neuron 35:625–641.
[7] Buzsaki G, Moser EI. 2013. Memory, navigation and theta rhythm in the hippocampal-entorhinal system. Nat Neurosci 16:130–138.
[8] McNamara RK, Skelton RW. 1993. The neuropharmacological and neurochemical basis of place learning in the Morris water maze. Brain Res Rev 18:33–49.
[9] Moser EI, Krobert KA, Moser M-B, Morris RGM. 1998. Impaired spatial learning after saturation of long-term potentiation. Science 281:2038–2042.
[10] Penner MR, Mizumori SJ. 2012. Neural systems analysis of decision making during goal-directed navigation. Prog Neurobiol 96:96–135.
[11] Suh J, Rivest AJ, Nakashiba T, Tominaga T, Tonegawa S. 2011. Entorhinal cortex layer III input to the hippocampus is crucial for temporal association memory. Science 334:1415–1420
[12] Whitlock JR, Heynen AJ, Shuler MG, Bear MF. 2006. Learning induces long-term potentiation in the hippocampus. Science 313:1093–1097
[13] Hollup SA, Molden S, Donnett JG, Moser M-B, Moser EI. 2001. Accumulation of hippocampal place fields at the goal location in an annular watermaze task. J Neurosci 21:1635–1644.
[14] Morris R. 1984. Developments of a water-maze procedure for studying spatial learning in the rat. J Neurosci Methods 11:47–60.
[15] Stewart CA, Morris RGM. 1993. The watermaze. In: Sahgal A, ed.Behavioural Neuroscience, Volume I, A Practical Approach. Oxford: IRL Press at Oxford University Press. p 107–122
[16] Morris RGM. 1981. Spatial localization does not require the presence of local cues. Learn Motiv 12:239–260
[17] Morris R. 1984. Developments of a water-maze procedure for studying spatial learning in the rat. J Neurosci Methods 11:47–60.
[18] Stewart CA, Morris RGM. 1993. The watermaze. In: Sahgal A, ed.Behavioural Neuroscience, Volume I, A Practical Approach. Oxford: IRL Press at Oxford University Press. p 107–122
[19] Vorhees CV, Williams MT. 2006. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat Protocols 1:848–858
[20] Wenk GL. 2004. Assessment of spatial memory using the radial arm and Morris water maze. In: Crawley JN, Gerhardt GA, Rogawski MA, Sibley DR, Skolnick P, Wray S, eds. Current Protocols in Neuroscience. New York: Wiley Interscience. Unit 8.5A
[21] Mactutus CF, Booze RM. 1994. Accuracy of spatial navigation: The role of platform and tank size. Soc Neurosci Abst 20:1014
[22] Williams MT, Braun AA, Amos-Kroohs R, McAllister JPI, Lindquist DM, Mangao FT, Vorhees CV, Yuan W. 2014. Kaolin-induced ventriculomegaly at weaning produces long-term learnng, memory, and motor deficits in rats. Int J Dev Neurosci 35:7–15.
[23] Schaefer TL, Lingrel JB, Moseley AE, Vorhees CV, Williams MT. 2011. Targeted mutations in the Na,K-ATPase alpha 2 isoform confer ouabain resistance and result in abnormal behavior in mice. Synapse 65:520–531
[24] Schaefer TL, Lingrel JB, Moseley AE, Vorhees CV, Williams MT. 2011. Targeted mutations in the Na,K-ATPase alpha 2 isoform confer ouabain resistance and result in abnormal behavior in mice. Synapse 65:520–531
[25] Williams MT, Morford LL, Wood SL, Wallace TL, Fukumura M, Broening HW, Vorhees CV. 2003b. Developmental d-methamphetamine treatment selectively induces spatial navigation impairments in reference memory in the Morris water maze while sparing working memory. Synapse 48:138–148.
[26] Williams MT, Moran MS, Vorhees CV. 2004. Behavioral and growth effects induced by low dose methamphetamine administration during the neonatal period in rats. Int J Dev Neurosci 22:273–283
[27] Vorhees CV, Herring NR, Schaefer TL, Grace CE, Skelton MR, Johnson HL, Williams MT. 2008. Effects of neonatal (+)-methamphetamine on path integration and spatial learning in rats: effects of dose and rearing conditions. Int J Dev Neurosci 26:599–610.
[28] Maurer R, Derivaz V. 2000. Rats in a transparent morris water maze use elemental and configural geometry of landmarks as well as distance to the pool wall. Spatial Cogn Comput 2:135–156
[29] Morris RGM, Schenk F, Jarrard LE. 1990. Ibotenate lesions of hippocampus and/or subiculum: Dissociating components of allocentric spatial learning. Eur J Neurosci 2:1016–1028
[30] Hamilton DA, Akers KG, Weisend MP, Sutherland RJ. 2007. How do room and apparatus cues control navigation in the Morris water task? Evidence for distinct contributions to a movement vector. J Exp Psychol Anim Behav Process 33:100–114
[31] Morris RGM. 1981. Spatial localization does not require the presence of local cues. Learn Motiv 12:239–260
[32] Vorhees CV, Reed TM, Acuff-Smith KD, Schilling MA, Cappon GD, Fisher JE, Pu C. 1995. Long-term learning deficits and changes in unlearned behaviors following in utero exposure to multiple daily doses of cocaine during different exposure periods and maternal plasma cocaine concentrations. Neurotoxicol Teratol 17:253–264
[33] Vorhees CV, Williams MT. 2006. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat Protocols 1:848–858
[34] Whishaw IQ, Tomie J-A. 1997. Perseveration on place reversals in spatial swimming pool tasks: Further evidence for place learning in hippocampal rats. Hippocampus 7:361–370
[35] Hoh T, Beiko J, Boon F,Weiss S, Cain DP. 1999. Complex behavioral strategy and reversal learning in the water maze without NMDA receptor dependent long-term potentiation. J Neurosci 19:RC2
[36] Baldi E, Efoudebe M, Lorenzini CA, Bucherelli C. 2005. Spatial navigation in the Morris water maze: working and long lasting reference memories. Neurosci Lett 378:176–180
[37] Rauch TM, Welch DI, Gallego L. 1989a. Hyperthermia impairs retrieval of an overtrained spatial task in the Morris water maze. Behav Neural Biol 52:321–330
[38] Rauch TM,Welch DI, Gallego L. 1989b. Hypothermia impairs performance in the Morris water maze. Physiol Behav 45:315–320
[39] Commins S, Cunningham L, Harvey D, Walsh D. 2003. Massed but not spaced training impairs spatial memory. Behav Brain Res 139:215–223
[40] Hamilton DA, Akers KG, Weisend MP, Sutherland RJ. 2007. How do room and apparatus cues control navigation in the Morris water task? Evidence for distinct contributions to a movement vector. J Exp Psychol Anim Behav Process 33:100–114
[41] Vorhees CV, Herring NR, Schaefer TL, Grace CE, Skelton MR, Johnson HL, Williams MT. 2008. Effects of neonatal (+)-methamphetamine on path integration and spatial learning in rats: effects of dose and rearing conditions. Int J Dev Neurosci 26:599–610.
[42] Vorhees CV, Herring NR, Schaefer TL, Grace CE, Skelton MR, Johnson HL, Williams MT. 2008. Effects of neonatal (+)-methamphetamine on path integration and spatial learning in rats: effects of dose and rearing conditions. Int J Dev Neurosci 26:599–610.
[43] Jonasson Z. 2005. Meta-analysis of sex differences in rodent models of learning and memory: A review of behavioral and biological data. Neurosci Biobehav Rev 28:811–825
[44] Tees RC, Buhrmann K, Hanley J. 1990. The effect of early experience on water maze spatial learning and memory in rats. Dev Psychobiol 23:427–439
[45] Vorhees CV, Herring NR, Schaefer TL, Grace CE, Skelton MR, Johnson HL, Williams MT. 2008. Effects of neonatal (+)-methamphetamine on path integration and spatial learning in rats: effects of dose and rearing conditions. Int J Dev Neurosci 26:599–610.