
While much of the research in behavioral sciences has focused primarily, often exclusively, on male subjects, a recent renewed focus on sexually divergent behavioral phenotypes has unveiled a plethora of differences between the sexes. Researchers should consider that outcome measures should be tailored to the group which they are investigating and that the magnitude of certain effects will be different in male and female rats and mice. Furthermore, male and female rats exhibit differences in the long-term impacts of behavioral interventions including stress. On a shorter time-scale, female rats exhibit different behavioral profiles throughout different points of their estrus cycle, another point which must be considered by researchers when performing research on both sexes. Here, we will discuss these points further including several examples of these differences.
Learning and memory
With regards to tests of learning and memory, a recent meta-analysis has suggested that male rodents have an overall higher performance across multiple tests of spatial learning.[1] For the Morris water maze, for instance, an analysis of nearly twenty studies from across two decades shows that on average, male mice and rats appear to learn the task faster than their female counterparts. This effect is independent on age and appears to be made more robust by pre-training exposures to the arena, further indicating a male-dominant paradigm for spatial learning. Interestingly, this apparent advantage for males in spatial learning and memory also extends to other tests, including the radial arm maze. The inclusion or exclusion of reward or external visual cues in these tests does not appear to impact this sex-difference, indicating that there is an inherent advantage of male rodents in tasks relying on spatial navigation and memory. It is therefore important that researchers perform their tasks of this nature in both sexes and compare any treatment conditions both within and between the male and female groups in order to best understand their results.
Aside from the sexually dimorphic results seen in tests of spatial memory, there are many other common tasks for which researchers should expect to see differences in the outcome measures between male and female rodents. Importantly, these differences are not always a matter of magnitude (i.e. spatial learning acquisition rates) but may also be distinct behaviors performed by one sex or another. For example, the classical approach to fear conditioning used to study stress, anxiety, and such conditions as post-traumatic stress disorder (PTSD) is to repeatedly pair an aversive stimulus (such as a footshock) with a previously neutral event (such as a tone). Over time, the tone (which comes to be known as the conditioned stimulus) becomes associated with the footshock (the unconditioned stimulus) and elicits a behavioral response (the conditioned response).[2] As this test was developed in male rats, the common conditioned response is for the animal to freeze. Thus, studies which use fear conditioning as a model are likely to display their results as a function of the time spent freezing, which increases with repeated pairings and decreases as the conditioned stimulus is repeatedly presented with non-aversive stimuli or is presented alone (a process known as “extinction”). Female rats, when presented with this same task, do not exhibit similar levels of freezing to their male counterparts. Rather, female rats make an abrupt, quick movement known as “darting” as a response to the conditioned stimulus.[3] This “darting” is similarly increased over repeated pairings of
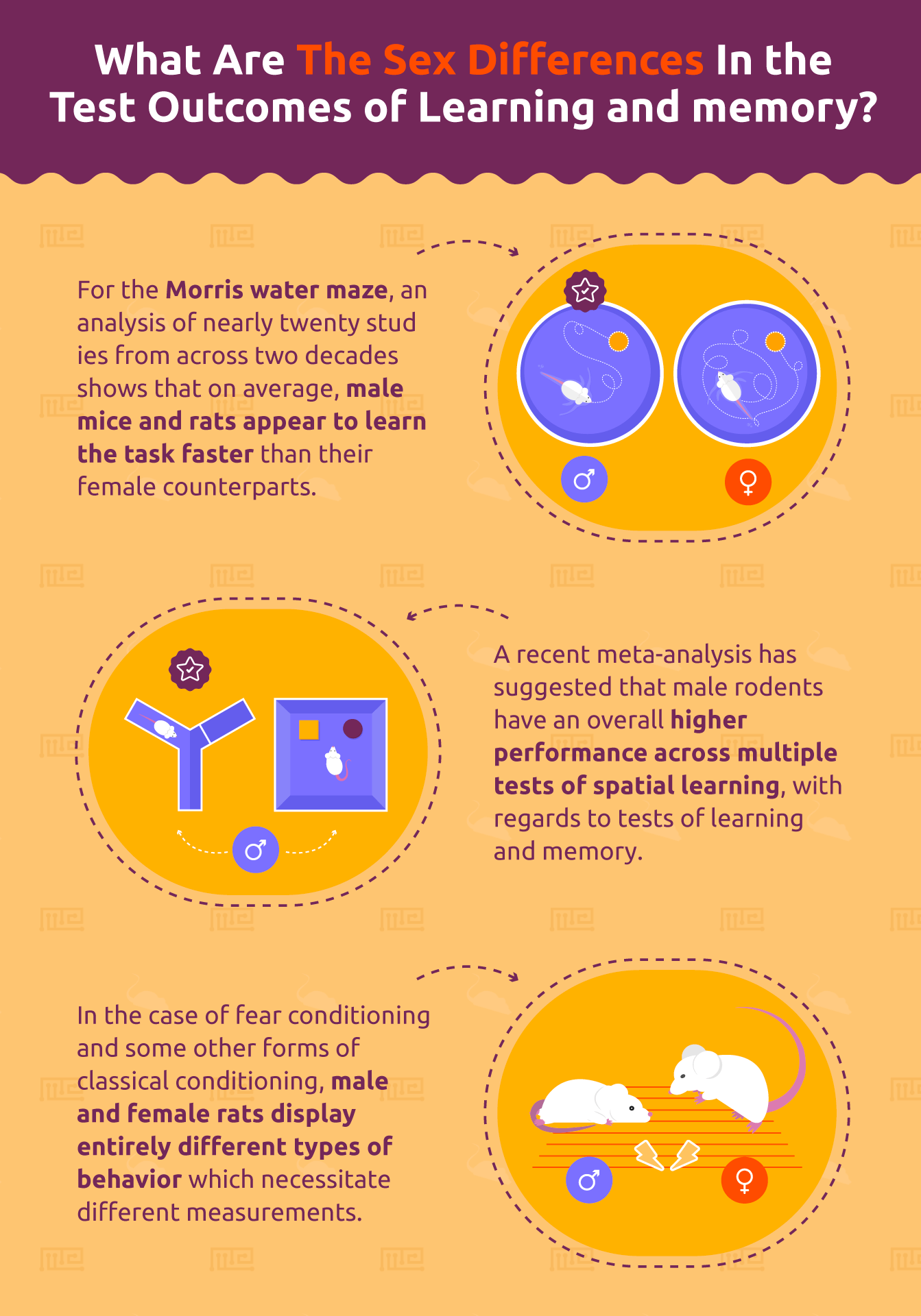
the conditioned and unconditioned stimuli and similarly decreases with extinction procedures, suggesting that behaviorally it is an equivalent measure to freezing behavior. Thus, in the case of fear conditioning and some other forms of classical conditioning,[4] male and female rats display entirely different types of behavior which necessitate different measurements.
Changes during the estrus cycle
An additional important consideration for the analysis of behavioral tests regarding sexual divergence is the estrus cycle. Whether researchers use ovariectomized or intact female rats, levels of estrogen fluctuate during the estrus cycle. It has been shown that the level of estrogen has significant effects on multiple behaviors, spanning from learning paradigms to baseline anxiety. For instance, compulsive lever-pressing fluctuates during the estrus cycle,[5] indicating that tests which use operant chambers may have different outcomes in female rodents during diestrus (where compulsive behaviors are highest) as compared to proestrus (where they are lowest). Furthermore, the period of proestrus alters performance in a variety of cognitive tasks. Novel object recognition and spontaneous alternation in the T-maze task are both improved during proestrus and performance decreases during diestrus.[6] Anxiety-like behaviors are also different when compared between these two periods. Female rats spend more time in the open arms of an elevated plus maze during proestrus as well as more time in the mirror in the mirror chamber task, both indicated reduced levels of anxiety as compared to during diestrus. Interestingly, this proestrus/diestrus difference was not seen in the open field task, suggesting that it is not a universally applicable phenomenon. In light of these findings, researchers should take care to note the differences between sexes with regards to behavioral performance and consider additionally, the influence of estrus cycle phases when interpreting their behavioral measures. Comparing female rats or mice within group (i.e. against other female rats/mice) must be done with animals who were tested during the same period of the estrous cycle in order to avoid errant behavioral differences which may be related to this flux rather than any other behavioral intervention being tested (for more information on this, see: “Does Estrous Cycle Affect Behavioral Testing Results In Mice?”).
Protracted sex-differences
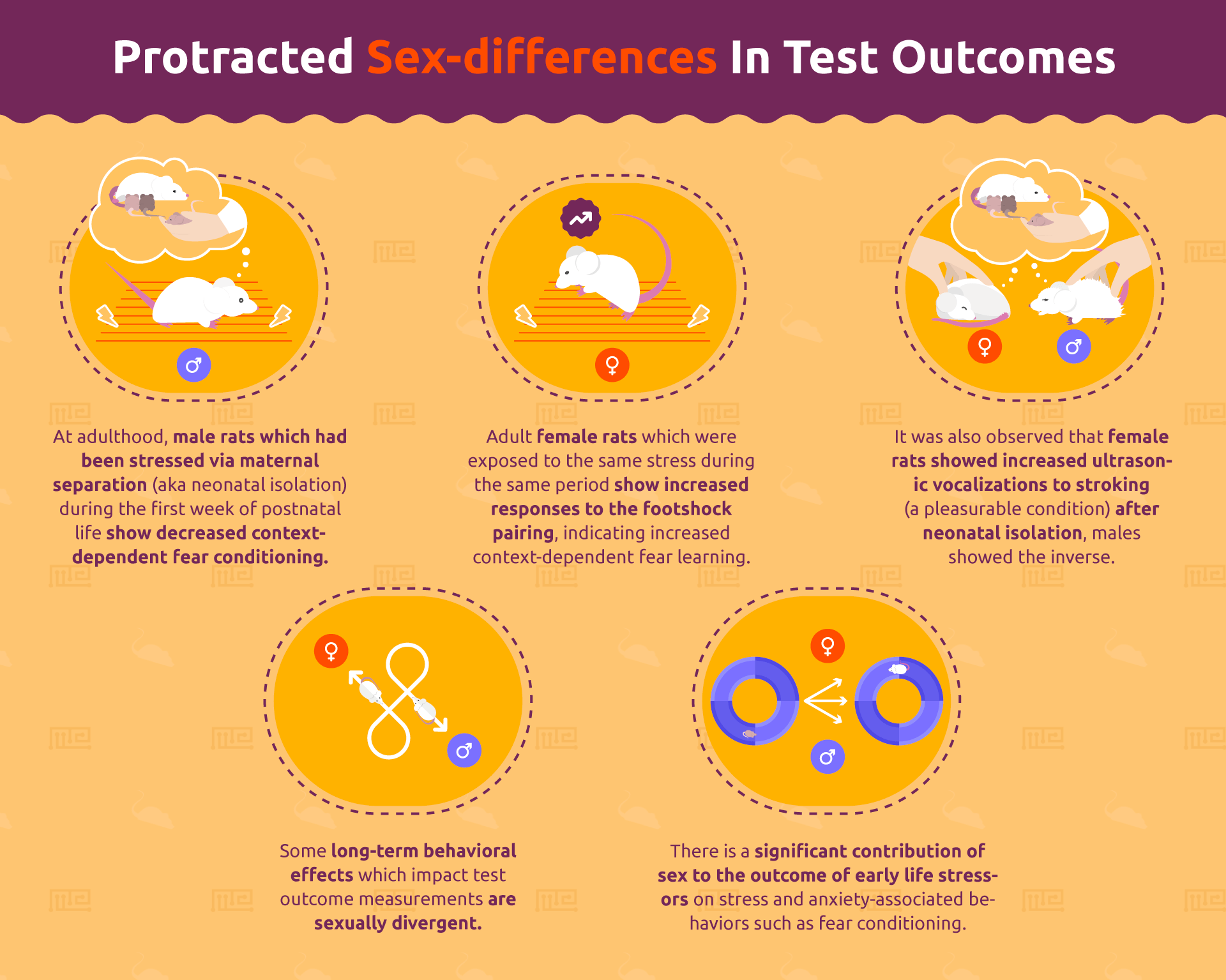
Finally, it should be noted that some long-term behavioral effects which impact test outcome measurements are also sexually divergent. For instance, neonatal isolation is a common means of inducing early-life stress in developing rodents. Rats which have been exposed to this early-life stressor exhibit altered behavior later in life, which manifests differently in males and females. At adulthood, male rats which had been stressed via maternal separation (aka neonatal isolation) during the first week of postnatal life show decreased context-dependent fear conditioning.[7] On the other hand, adult female rats which were exposed to the same stress during the same period show increased responses to the footshock pairing, indicating increased context-dependent fear learning. Some debate exists as to whether or not this effect is similarly manifested in situations of appetitive, rather than aversive, behaviors. While the above researchers also found that female rats showed increased ultrasonic vocalizations to stroking (a pleasurable condition) after neonatal isolation, males showed the inverse. However, context-conditioning with the opioidergic drug morphine does not appear to be impacted by these procedures in a sex-dependent manner.[8] Nonetheless, it is clear that there is a significant contribution of sex to the outcome of early life stressors on stress and anxiety-associated behaviors such as fear conditioning.
Conclusions
Clearly, researchers should be aware that there are significant differences between male and female rodents in multiple respects regarding behavioral testing. First, the magnitude of certain outcome measures will be different between the two sexes, such as in spatial learning and memory tasks. Additionally, there are distinct behaviors that are displayed by males but not females, or vice versa. Examples include play behaviors[9] and fear-responding (e.g. freezing versus darting in response to a conditioned stimulus). Finally, certain long-term behavioral changes, such as altered fear-responding as a result of early-life stress, are inversely expressed in males and females. Thus, it is paramount that researchers identify such potential differences between male and female rodents prior to starting their experiments in order to ensure that they are collecting the appropriate measurements for their task and sex. Further, statistical analyses of behavioral outcome measures should always be done both within and between sexes in order to identify such differences.
References
- Jonasson, Z. (2005). Meta-analysis of sex differences in rodent models of learning and memory: a review of behavioral and biological data. Neuroscience & Biobehavioral Reviews, 28(8), 811–825.
- Blanchard, R. J., & Blanchard, D. C. (1969). Passive and active reactions to fear-eliciting stimuli. Journal of Comparative and Physiological Psychology, 68(1), 129–135.
- Gruene, T. M., Flick, K., Stefano, A., Shea, S. D., & Shansky, R. M. (2015). Sexually divergent expression of active and passive conditioned fear responses in rats. eLife, 4, e11352.
- Dalla, C., & Shors, T. J. (2009). Sex differences in learning processes of classical and operant conditioning. Physiology & behavior, 97(2), 229-38.
- Flaisher-Grinberg, S., Albelda, N., Gitter, L., Weltman, K., Arad, M., & Joel, D. (2009). Ovarian hormones modulate “compulsive” lever-pressing in female rats. Hormones and Behavior, 55(2), 356–365.
- Walf, A. A., Koonce, C., Manley, K., & Frye, C. A. (2008). Proestrous compared to diestrous wildtype, but not estrogen receptor beta knockout, mice have better performance in the spontaneous alternation and object recognition tasks and reduced anxiety-like behavior in the elevated plus and mirror maze. Behavioural brain research, 196(2), 254-60.
- Kosten, T. A., Miserendino, M. J. D., Bombace, J. C., Lee, H. J., & Kim, J. J. (2005). Sex-selective effects of neonatal isolation on fear conditioning and foot shock sensitivity. Behavioural Brain Research, 157(2), 235–244.
- Knutson B, Burgdorf J, Pankseep J. High-frequency ultrasonic vocalizations index conditioned pharmacological reward in rats. Physiol and Behav 1999;66:639–43.
- Meaney MJ, Stewart J, Beatty WW. Sex-differences in social play – the socialization of sex-roles. Advances in the Study of Behavior. 1985;15:1–58.